10.1.1. Облегчение
Изменение
эффективности синаптической передачи,
происходящее при высокой активности
синапса, можно пронаблюдать в следующем
опыте. Представим себе, что мы осуществляем
микроэлектродное отведение от области
частично кураризованной концевой
пластинки скелетной мышцы лягушки и
подаем на двигательное волокно два
стимула, причем второй из этих стимулов
следует за первым через различные
промежутки времени (рис. 6–42). Если второй
постсинаптический потенциал возникает
до окончания первого, то эти потенциалы
будут, естественно, суммироваться;
однако амплитуда этого второго потенциала
будет больше, чем должно было бы быть в
результате простой суммации. Если второй
постсинаптический потенциал возникает
вскоре после окончания первого, его
величина будет больше, чем первого. В
двигательной концевой пластинке это
явление, получившее название синаптического
облегчения,длится
100–200 мс.
|
Рис. |
По
имеющимся данным облегчение обусловлено
скорее всего тем, что поступление в
пресинаптическое волокно первого
импульса сопровождается повышением в
нем концентрации свободных ионов Са2+ ,
которое сохраняется непродолжительное
время. В результате, когда в волокно
поступает второй импульс и тоже вызывает
увеличение i,
эта новая «порция» ионов Са2+ суммируется
с тем количеством этих ионов, которое
осталось от первого возбуждения.
Поскольку же выделение медиатора связано
степенной зависимостью с внутриклеточной
концентрацией Са2+ в
области пресинаптических участков
высвобождения этого медиатора (разд.
6.8.2), небольшое повышение i приводит
к существенному увеличению количества
медиатора, высвобождаемого в ответ на
второй импульс. Экспериментальное
подтверждение этой гипотезы было
получено Бернардом Катцем и Рикардо
Миледи (1968). Эти исследователи подводили
микроэлектрод как можно ближе к области
двигательной концевой пластинки мышцы
лягушки, погруженной в бескальциевый
раствор Рингера, и через этот микроэлектрод
посылали «залпы» Са2+ (рис.
6–43, А).Оказалось,
что ПКП возникает лишь тогда, когда ко
времени прихода в пресинаптическое
окончание импульса во внеклеточной
среде имеются ионы Са2+ (рис.
6–43, Б,
В).Катц
и Миледи обнаружилитакже, что облегчение
постсинаптического потенциала в ответ
на второй стимул было более выраженным,
если Са2+ подавался
из микроэлектрода во внеклеточную среду
одновременно с поступлением первого
ПД (рис. 6–43, Д).Если
первый кальциевый залп подавался после
поступления первого импульса в окончание,
существенного облегчения не наблюдалось
(рис. 6–43, Г).Значит,
для того чтобы произошло облегчение, к
моменту прихода импульса в пресинаптическое
волокно в среде должен содержаться
кальций, который может войти в это
волокно. Как мы уже говорили, по–видимому,
некоторая часть ионов Са2+,
вошедших в волокно при возбуждении,
остается в нем и суммируется с тем
количеством кальция, который поступает
в ответ на второй импульс. В результате
концентрация Са2+ в
волокне увеличивается, высвобождается
больше медиатора, а следовательно,
повышается постсинаптический потенциал.
|
Рис. |
Синтез медиатора
Медиаторы
(«посредники») обеспечивают одностороннюю
передачу возбуждения — от нервного
волокна к эффекторной клетке рабочего
органа или к другому нейрону. Некоторые
медиаторы (например, ацетилхолин)
синтезируются в цитоплазме синаптических
окончаний из нормальных компонентов
внутри- и внеклеточных жидкостей,
поступающих в них путем ниноцитоза.
Другие образуются в теле нейрона, в
частности в аппарате Гольджи, а затем
путем прямого аксонного транспорта
поступают в синаптическое окончание и
депонируются в везикулах.
Путем
постоянного медленного аксонного
транспорта от тела нейрона к окончаниям
аксона движутся белки и ферменты,
участвующие в синтезе медиатора в
пресинаптической бляшке. Быстрый
антероградный транспорт доставляет- в
синаптическое окончание пузырьки с
готовым медиатором.
В
синапсах скелетных мышц «посредником»
является только один вид медиатора —
ацетилхолин. Он синтезируется из холина
(продукт печеночной секреции), уксусной
кислоты и фермента коэнзима-А, регулирующего
активность метаболических процессов.
Синапсы, в которых медиатором является
ацетилхолин, называются холинергическими.
Ацетилхолин активирует натриевые ионные
каналы в цитоплазматических мембранах
исчерченных мышечных волокон, способствуя
развитию потенциала действия, активации
кальциевых каналов и сокращению скелетных
мышц.
В
синапсах внутренних органов и сосудов
функции медиатора выполняет и норадреналин,
относящийся, как и адреналин, к
катехолсеминам. Он синтезируется из
аминокислот тирозина и фенилаланина.
Синапсы, в которых медиатором является
норадреналин, называются адренергическими.
Варикозные
расширения синапсов гладких мышц
содержат как ацетилхолин, так и
норадреналин, которые могут оказывать
различное физиологическое воздействие,
в зависимости от вида активируемых ими
рецепторов и каналов.
В
синапсах мозга функции медиаторов
выполняют более 30 биологически активных
веществ. К наиболее изученным медиаторам
относятся норадреналин, ацетилхолин,
некоторые моноамины (адреналин, серотонин,
дофамин). А также аминокислоты (глицин,
глутаминовая кислота, гаммааминомасляная
кислота); нейропептиды — энкефалины,
нейротензин, ангиотензин II и др.
Медиаторы
синтезируются постоянно и депонируются
в везикулах (пузырьки) синаптических
окончаний.
Диффузии медиатора через постоянные поры.
Постсинаптическая
мембрана синапса.
В постсинаптических
мембранах имеются 2 типа рецепторов:
ионотропные и метаботропные.
Ионотропные
рецепторы постсинаптической мембраны.
При воздействии
медиатора на ионотропные рецепторы
постсинаптической мембраны развиваются
быстрые реакции, длящиеся несколько
миллисекунд.
При этом возникают
реакции двух типов:
ВПСП — возбуждающий
постсинаптический потенциал.
ТПСП – тормозной
постсинаптический потенциал.
ВПСП (возбуждающий
постсинаптический потенциал) возникает
при взаимодействии медиатора с
ионотропными рецепторами, ассоциированными
с натриевыми ионными каналами за
счет развития деполяризации
постсинаптической мембраны,связанной
с формированием входящего натриевого
тока.
ВПСП хорошо
распространяется в пределах одного
нейрона.
ТПСП (тормозной
постсинаптический потенциал) возникает
при взаимодействии медиатора с
ионотропными рецепторами, ассоциированными
с калиевыми или хлорными ионными
каналами, за счет развития гиперполяризации
постсинаптической мембраны, связанной
либо с усилением выходящего калиевого
тока, либо с увеличением входящего
тока хлора.
Метаботропные
рецепторы постсинаптической мембраны.
Метаботропные
рецепторы постсинаптической мембраны
при взаимодействии с медиатором
активируют преимущественно семейство
высокомолекулярных G-белков,
которые в свою очередь активируют
основные каскадные ферментные системы
(аденилатциклазную, фосфолипазную)
усиления и реализации информационного
сигнала в клетке.
Кроме того,
медиаторы, взаимодействуя с рецепторами
постсинаптической мембраны, способны
прямо или опосредованно активировать
образование цГМФ.
Активация
метаботропных рецепторов, вызывающая
сложные биохимические и, как следствие,
функциональные перестройки в клетке
(медленные реакции), часто прямо или
опосредованно сопряжена с изменением
проницаемости ионных каналов (быстрые
эффекты).
Это обусловлено
тем, что многие компоненты каскадных
систем реализации сигнала либо
ассоциировано (субъединица Gβγ),
либо конститутивно (протеинкиназы,
вторые посредники) способны менять
активность белков, формирующих ионные
каналы.
К большинству
медиаторов имеются и ионотропные,
и метаботропные рецепторы.
Система
удаления медиатора из синаптической
щели.
1. Система обратного
транспорта медиатора (описана в
разделе «область пресинаптической
мембраны»)
2. Ферменты,
метаболизирующие медиаторы (холинэстераза,
КОМТ, МАО и др.). Они метаболизируют
медиаторы, прекращая их действие на
рецептор.
Ко-медиаторы.
Ко-медиаторы это
дополнительные химические посредники
передачи информации, к которым на
постсинаптической мембране синапса
выделены специфические рецепторы.
Рецепторы к
ко-медиаторам относятся к метаботропным
рецепторам.
В области
пресинаптической мембраны ко-медиаторы
содержатся в крупных везикулах. Они
относятся к нейропептидам. В каждой
везикуле содержится несколько десятков
молекул ко-медиатора.
Особенностью
выделения ко-медиаторов из области
пресинаптической мембраны является
сопряженность их выделения с
классическими медиаторами.
Выделение
ко-медиатора из области пресинаптической
мембраны осуществляется путем
экзоцитоза.
Ко-медиаторы
видоизменияют ответ постсинаптического
нейрона на действие классических
медиаторов.
Нейромодуляторы.
Нейромодуляторы
по сравнению с нейромедиаторами
имеют ряд особенностей действия:
1. Нейромодуляторы
не обладают самостоятельным
физиологическим действием, а модифицируют
эффект нейромедиаторов.
2. Действие
нейромодуляторов имеет тонический
характер – медленное развитие и большую
продолжительность действия (секунды,
минуты).
3. Действие
нейромодуляторов не сопряжено во
времени с эффектом нейромедиатора.
В постсинаптической
мембране имеются метаботропные
рецепторы к нейромодуляторам,
которые обладают низкой специфичностью.
Для синапсов ЦНС
в качестве нейромодуляторов выступают
более десяти семейств нейропептидов.
В мозге несколько
нейромодуляторов могут действовать
на один тип синапсов и один нейромодулятор
— на несколько типов синапсов.
Как зависит амплитуда тетанического сокращения скелетной мышцы от частоты стимуляции нерва
Амплитуда
гладкого тетануса увеличивается при
увеличении частоты стимуляции нерва и
при некоторой оптимальной частоте
возникает оптимальный тетанус (оптимум)
Это связано с тем, что при увеличении
частоты ионизированный Са в пресинаптическом
окончании увеличивается, увеличивается
выброс медиатора , также возрастает
концентрация Са в зоне протофибрилл
,что улучшает взаимодействия актина и
миозина..
28.Как
доказать, что пессимум не связан с
утомлением мышцы?
Увеличение
частоты стимуляции сверх оптимума
приводит к расслаблению мышцы (пессимум)
.Если же частоту уменьшить, то мышца
вновь сокращается, что доказывает, что
пессимум не является утомлением.
Железы
внутренней секреции
1.Гормональная
регуляция физиологических функций.
Особенно
важную роль в гуморальном взаимодействии
органов, тканей и клеток играют те из
них, которые имеют специализированную
способность вырабатывать вещества,
изменяющие состояние организма, функцию
и структуру органов и тканей. Эти вещества
называют гормонами, а выделяющие их
органы — эндокринными железами или
железами внутренней секреции. Они
названы так потому, что в отличие от
желез внешней секреции не имеют выводных
протоков и выделяют образующиеся в них
вещества непосредственно в кровь.
К железам
внутренней секрецииотносятся гипофиз,
щитовидная железа, околощитовидные
железы, островковый аппарат поджелудочной
железы, кора и мозговое вещество
надпочечников, половые железы и плацента,
эпифиз.
Гормоны
обладают дистантным действием, поступая
в кровяное русло, они оказывают влияние
на весь организм, а также на органы и
ткани, расположенные вдали от той железы,
где они образуются.
Выделяют
четыре типа влияния гормонов на организм:
1) метаболическое
(действие на обмен веществ);
2) морфогенетическое
(стимуляция формообразовательных
процессов, дифферениировки, роста и
пр.);
3) кинетическое
(включающее определенную деятельность
исполнительных органов);
4) корригирующее
(изменяющее интенсивность функции
органов и тканей).
Характерным
свойством гормонов является их высокая
физиологическая активность. Это означает,
что очень малое количество гормона
может иызвать изменения функций
организма. Гормоны быстро разрушаются
в тканях, в частности в печени, поэтому
необходимо постоянное их выделение
соответствующей железой.
Известные
гормоны позвоночных могут быть разделены
на три основных класса:
1) стероиды;
2) производные
аминокислот;
3) белково-пептидные
соединения.
Регуляция
функций эндокринных желез осуществляется
несколькими способами:
1) прямое
влияние на клетки железы концентрации
в крови того вещества, уровень которого
регулирует данный гормон;
2) опосредованное,
нейрогуморальное влияние.
25.Опишите механизм сокращения скелетной мышцы. Одиночное сокращение и его фазы.
В состоянии
покоя актини миозин не взаимодействуют
друг с другом, так как их активные центры
закрыты. В покое активный центр актина
закрыт белком – тропомиозином, к которому
прикреплён белок – тропомин.
При возбуждении
ионы Са взаимодействуют с тропомином,
вызывая конформациютропомиозина.
Активный центр миозина открывается,
формируется поперечный мостик между
актином и миозином, эти мостики начинают
совершать гребковые движения, обеспечивая
скольжение актина вдоль миозина и
сокращение мышцы.
Одиночное
сокращениевозникает в ответ на
одиночный стимул или одиночный ПД.
Первый— латентный период представляет собой
сумму временных задержек, обусловленных
возбуждением мембраны мышечного
волок¬на, распространением ПД по
Т-системе внутрь волокна, образованием
инозитолтрифосфата, повышением
концентрации внутриклеточного кальция
и активации поперечных мостиков. Для
портняжной мышцы лягушки латентный
период составляет около 2 мс.
Второй— период сокращения. В случае свободного
укорочения мышечного волокна говорят
об изотоническом режиме сокращения,
при котором напряжение практически не
изменяется, а меняется только длина
мышечного во¬локна. Если мышечное
волокно закреплено с двух сторон и не
может свободно укорачиваться, то говорят
об изометрическом режиме со¬кращение.
Третий —
период расслабления, когда уменьшается
кон¬центрация ионов Са2+ и отсоединяются
головки миозина от актиновыхфиламентов.
Понятие и физиология химических и электрических синапсов
 |
Синапс — это специализированная структура, обеспечивающая межклеточную передачу сигналов электрической и (или) химической природы.
С помощью синапсов передается информация от рецепторных клеток на дендриты чувствительных нейронов, с одной нервной клетки на другую, с нервной клетки на волокно скелетной мышцы, железистые и другие эффекторные клетки. Через синапсы могут оказываться возбуждающие или тормозные влияния на клетки, активироваться или подавляться их метаболизм и другие функции.
Строение синапса
Электронно-микроскопические исследования выявили, что синапсы имеют три основных элемента: пресинаптическую мембрану, постсинаптическую мембрану и синаптическую щель (рис. 1).
Передача информации через синапс может осуществляться химическим или электрическим путем. Смешанные синапсы сочетают химические и электрические механизмы передачи.
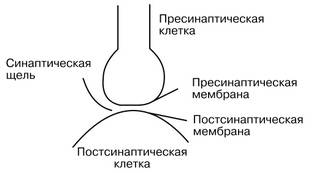
Рис. 1. Основные элементы синапса
Виды синапсов
По механизму передачи возбуждения синапсы подразделяют на электрические и химические.
Электрические синапсы образуются между клетками, формирующими между мембранами плотные щелевые контакты. Ширина щели составляет около 3 нм, и между контактирующими мембранами образуются общие ионные каналы с диаметром поры около 1-2 нм. Через эти каналы и осуществляется передача информации с помощью электрических ионных токов. Через каналы электрических синапсов клетки могут обмениваться также небольшими по размеру сигнальными молекулами органической природы. Названные вещества способны перемещаться в электрических синапсах с большой скоростью в обоих направлениях, и переносимая с их помощью информация также может передаваться в обоих направлениях (в отличие от химических синапсов).
Электрические синапсы имеются уже в эмбриональном мозге и остаются наряду с химическими синапсами в зрелой ЦНС позвоночных.
Ионные токи, перемещающиеся из пресинаптического нейрона в постсинаптический, вызывают на его мембране колебания разности потенциалов — постинаптический потенциал амплитудой около 1 мВ и могут вызвать генерацию на ней ПД. В свою очередь возникший ПД может вызвать обратный ток ионов через каналы щелевых контактов к пресинаптическому нейрону и становится источником модуляции разности потенциалов на его мембране. Нейрон может формировать щелевые контакты (электрические синапсы) с рядом других нейронов, поэтому практически одновременное протекание ионных токов между ними способствует синхронизации активности группы нервных клеток, связанных этими синапсами. Электрические синапсы чаще выявляются в областях мозга, в которых регистрируется высоко синхронизированная нейронная активность.
Как уже упоминалось ранее, ионные каналы щелевых контактов имеются не только между нервными, но и между глиальными клетками, между гладкими миоцитами, между кардиомиоцитами, между железистыми клетками.
Химические синапсы образуются специализированными структурами двух клеток в области их контакта (рис. 2). Одной из этих клеток, которую называют пресинаптической, обычно является нервная клетка, но ею может быть и специализированная чувствительная клетка иной природы (например, сенсоэпителиальная слуховая или вкусовая клетка, гломусные клетки аортального тельца). Пресинаптическая нервная клетка обычно формирует синапс на другой клетке с помощью мембраны нервного окончания (аксона). В этом случае окончание аксона называют пресинаптической, или аксонной, терминально.
Узкое щелевидное пространство, разделяющее пресинаптическую и постсинаптическую мембраны, называют синаптической щелью (см. рис. 2.). Таким образом, для химических синапсов общими структурными элементами являются пресинаптическая часть (нервное окончание и пресинаптическая мембрана), синаптическая щель, постсинаптическая часть (постсинаптическая мембрана).
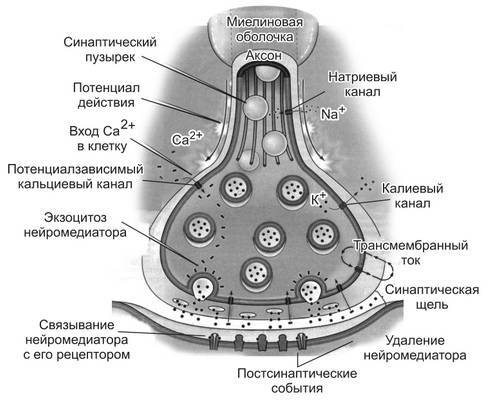
Рис. 2. Строение синапса и процессы, осуществляемые в ходе синаптической передачи сигнала
Химические синапсы могут образовываться между двумя нервными клетками с участием отростков и тела клетки. В зависимости от структур нейронов, образующих синаптическое соединение, синапсы делят на аксосоматические, аксоаксональные, аксодендритные, дендродендритные. Синапсы, располагающиеся в пределах ЦНС, называют центральными, а находящиеся вне ЦНС — периферическими. Периферические синапсы передают сигналы нервных волокон на эффекторные органы (мышечные волокна, железистые клетки).
Серотонин.
Синтез
серотонина (5-НТ)
идет в 2 стадии
1.
Из пищевой аминокислоты триптофана
образуется
5-гидрокситриптофан;
фермент
триптофангидроксилаза.
2.
Из 5-гидрокситриптофана образуется
5-гидрокситриптамин
(5-НТ; серотонин);
фермент
декарбоксилаза
ароматических аминокислот.
Серотонин
является тканевым гормоном (увеличивает
тонус гладких мышечных клеток в стенках
сосудов и ряда других внутренних
органов).
Кроме
того, серотонин – медиатор ЦНС;
вырабатывают нейроны ядер
шва
(верхне-центральная зона среднего мозга,
моста и продолговатого мозга с переходом
в спинной мозг); аксоны клеток ядер шва
расходятся по всей ЦНС, образуя контакты
обычного и варикозного типа.
Жизненный
цикл 5-НТ
-
В
верхней части рисунка – пресинаптическое
окончание. -
Синтез
5-НТ из триптофана. -
Перенос
5-НТ в везикулу. -
Экзоцитоз
5-НТ. -
Синаптическая
щель. -
Взаимодействие
5-НТ с постсинаптическим рецептором. -
Отросток
постсинаптической клетки. -
5-НТ
влияет на пресинаптический рецептор
(аутоторможение экзоцитоза). -
Обратный
захват 5-НТ. -
5-НТ
повторно загружается в везикулу либо
разрушается с помощью МАО. -
Продукт
распада 5-НТ гидрокси-индо-лил-уксусная
кислота удаляется из пресинаптического
окончания.
Рецепторы
5-НТ:
7
типов с подтипами (5-НТ1 …
5-НТ7);постсинаптические, кроме 5-НТ1В и
5-НТ1D
(пресинаптические);5-НТ1: тормозят
аденилатциклазу и экзоцитоз медиаторов
(ослабляя
выброс Glu
вызывают торможение; ослабляя выброс
ГАМК – возбуждение).
5-НТ2: активируют фосфолипазу С (фермент,
который через синтез ВтП вызывает
закрывание К+-каналов);5-НТ3:
ионотропные, имеют Na+-канал
(много
в гиппокампе);
5-НТ4
– 5-НТ7: активируют аденилатциклазу,
распространены существенно меньше (в
основном в базальных ганглиях и коре
больших п/ш).
Периферические
эффекты 5-НТ:
Серотонин
как тканевой гормон в наибольшем
количестве выделяется тромбоцитами:
—
повреждение стенок сосуда;
—
далее: активация тромбоцитов;
—
запуск реакций свертывания: превращение
белка плазмы фибриногена в фибрин
(фибриновая сеть – основа тромба);
—
параллельно из тромбоцитов выделяется
5-НТ, вызывающий сокращение гладких
мы-шечных клеток в стенках сосудов
(спазм сосудов уменьшает кровотечение).
Сбой
в работе этой системе может вести к
мигреням
(5% мужчин, 15% женщин).
Дело
в том, что в крови постоянно происходит
спонтанное разрушение тромбоцитов,
определяющее «фоновый» уровень 5-НТ
плазмы и во многом – тонус сосудов.
Иногда (по не очень понятной причине)
тромбоциты разрушаются активнее, чем
обычно. Это вызывает сначала рост тонуса
сосудов (в плазме слишком много 5-НТ), а
затем – их избы-
точное
расслабление (снижение содержания 5-НТ
плазмы из-за уменьшения числа разрушающихся
тромбоцитов). К падению 5-НТ в плазме
особенно чувствительны сосуды головного
мозга, они расширяются, возникает отек
и головная боль. Лечение – агонисты
5-НТ1-рецепторов (например, суматриптан).
3.
Миндалина.
Миндалина
относится к базальным ганглиям больших
полушарий; вместе с гипоталамусом
отвечает за многие биологические
потребности: пищевую, питьевую, половую
и родительскую, в безопасности (центры
страха и агрессии).
Центры
пищевой и питьевой потребностей (голода
и жажды) находятся в средней части
гипоталамуса и в меньшей мере связаны
с миндалиной.
Центры
полового и родит. поведения (передняя
часть гипоталамуса) работают вместе
с
миндалиной, «откликаясь» на изменения
концентрации ряда гормонов.
Центры
страха и агрессии (задняя часть
гипоталамуса) работают
под
управлением миндалины.
С
миндалиной также связана потребность
доминирования в стае и ряд других
«зоосоциальных» (по
П.В. Симонову)
потребностей.
При
на рушении миндалины наблюдается
нарушение процедуры смены и выбора
доминанты, нервная система может
«зависать» на той или иной потребности(мании,
психозы). При этом более «тонкие»
потребности нередко вообще исчезают(например,
стремление доминировать в стае).
Доминанту.
Именно
за счёт своей пластичности синапсы
могут «учиться» и переходить в
более возбуждённое состояние (сенситизация)
или в более заторможенное (привыкание).
Пространственная
и временная суммация основана на свойстве
каждого нейрона в центре к суммации как
возбуждения, так и торможения.
Поскольку каждый нервный центр имеет
много параллельно расположенных
афферентных или входных волокон от
рецептивного поля рефлекса, слабые
раздражения нескольких участков
рецептивного поля, в отдельности не
способные реализовать рефлекс, вызывают
в нейронах центра несколько ВПСП, которые
суммируются, приводя к формированию на
мембране нервной клетки потенциалов
действия, распространяющихся по
эфферентным проводникам, вызывая
рефлекторную реакцию. Это явление
называют пространственной
суммацией. При
увеличении частоты афферентных сигналов
в единицу времени амплитуда ВПСП
нарастает до критического уровня из-за
повышения эффективности синаптического
проведения, что также вызывает возбуждение
нейронов и рефлекторный ответ на слабые
частые раздражения. Это явление
называют временной
суммацией;
Пластичность нервных центров —
способность перестраивать функциональные
свойства для более эффективной регуляции
функций, осуществления
новых, ранее несвойственных
этому центру рефлексов
или восстановления функций
после повреждения
части нейронов центра. Пластичность
обеспечивает изменение эффективности
и направленности связей между нервными
клетками, является рабочим механизмом
обучения. В основе пластичности
лежат функциональные особенности
синапсов и мембран нейронов («тренировка»
синапсов, посттетаническая
потенциация, периодичность
функционирования синапсов и нейронов,
пространственная и временная суммация
постсинаптических потенциалов), а также
наличие многочисленных дублирующих
систем нейронов и нервных волокон;
Совершенно
очевидно, что можно выделить две группы
синаптических механизмов, ответственные
за изменения эффективности синаптической
передачи: пресинаптические и
постсинаптические. В первом случае
эффективность синаптической передачи
может зависеть от количества медиатора,
высвобождаемого пресинаптическими
окончаниями в ответ на поступление в
них ПД. Во втором случае могут происходить
какие–то изменения постсинаптических
структур, при которых меняется степень
деполяризации в ответ на действие
определенного количества медиатора. О
постсинаптической пластичности известно
сравнительно мало, хотя в некоторых
тканях она и была обнаружена. Поэтому
мы ограничимся лишь обсуждением
пресинаптических механизмов пластичности.
Существуют
две основные группы пресинаптических
механизмов, которые могут приводить к
изменениям эффективности синаптической
передачи. В первом случае изменения
свойств пресинаптического окончания
возникают в связи с его собственной
активностью, если эта активность высока;
подобные изменения обычно бывают
кратковременными. Такие механизмы носят
общее название гомосинаптической
модуляции. Во втором случае свойства
пресинаптических окончаний изменяются
под действием модулятора, высвобождаемого
другим, тесно прилегающим нервным
окончанием. Подобные сдвиги обычно
бывают более долговременными. Механизмы
такого рода называются гетеросинаптической
модуляцией.
II. Синаптические везикулы.
В большинстве
синапсов имеются 2 типа синаптических
везикул: мелкие и крупные.
Мелкие синаптические
везикулы содержат медиатор. В
одной везикуле содержится несколько
тысяч молекул медиатора, что
составляет квант медиатора.
Большая часть
мелких синаптических везикул
фиксирована к цитоскелету терминали.
Меньшая часть
синаптических везикул связана с
внутренней поверхностью пресинаптической
мембраны. Эти везикулы находятся
в активной зоне, поставляют медиатор
для очередного выброса.
Существует
постоянный переход мелких синаптических
везикул из первого пула во второй.
Образование
оболочек мелких везикул, ферментов,
необходимых для образования медиатора,
происходит в эндоплазматическом
ретикулуме и цистернах аппарата Гольджи,
после чего они аксоплазматическим
током транспортируются в область
пресинаптической мембране, где происходит
«упаковка» молекул медиаторов в везикулы.
Образование
медиатора идет непосредственно в
области пресинаптической мембраны.
«Упаковка»
медиатора в мелкие везикулы является
активным процессом.
Крупные
синаптические везикулы. В них содержатся
нейропептиды, которые играют в
синапсе роль нейромодуляторов, то
есть веществ, модифицирующих (ослабляющих,
усиливающих, тормозящих, потенцирующих
и др.) действие медиаторов.
Образование
крупных везикул происходитв
эндоплазматическом ретикулуме и
цистернах аппарата Гольджи, здесь же
происходит заключительный этап
формирования нейропептидов в
нейромодуляторы и «упаковка» их
в крупные везикулы.
После этого они
аксоплазматическим током транспортируются
в область пресинаптической мембраны.
Синтез медиаторов
и образование мелких и крупных
синаптических пузырьков осуществляется
непрерывно.
Механизм
выделения медиатора в пресинаптическую
щель.
Деполяризация
области пресинаптической мембраны,
которая связана с распространением
возбуждения по аксону, вызывает активацию
потенциалзависимых кальциевых каналов.
В области активной
зоны локально возникает кальциевый
домен, который активирует систему
белков экзоцитоза везикулярной и
пресинаптической мембран.
Активация белков
экзоцитоза приводит к выделению медиатора
в синаптическую щель по трем вариантам.
Путем:
Список источников
- StudFiles.net
- www.grandars.ru